
The oxidation of dopamine produces dopamine o-quinone, a melanin precursor. This reaction is mediated by Tyrosinase, an enzyme that catalyzes the oxidation of o-diphenols, such as dopamine, into o-quinoles using dioxygen. In pathological conditions, a reactive oxygen species is generated, namely dopamine o-semiquinone, and its formation is catalyzed by lactoperoxidase and hydrogen peroxide. Pathological conditions may involve the enzymatic breakdown of monoamine oxidases (MAOs) and/or the induction of oxidation reactions from environmental toxic agents. Monoamine oxidases normally recycle extra amounts of dopamine in the brain. The end product of the oxidation of dopamine is an Aminochrome that carries a detrimental function in parkinson's disease as it has been shown to damage essential cellular macromolecules and increase the production of ROS.

Briefly, Aminochrome undergoes a one-electron reduction reaction where redox cycling DA-semiquinones are formed. The enzyme that mediates the reduction of Aminochrome to the o-semiquinone is the NAPDH cytochrome P450 reductase. Dioxygen species are oxidized to ROS while NADPH to NADP. Interestingly, antioxidants such as Superoxide Dismutase and catalase exacerbate this reaction since it depletes the cell fr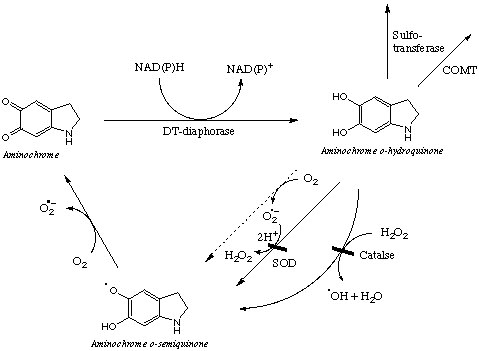 om superoxide species, thus moving the reaction towards the reactant (aminochrome). Thus, the presence of SOD and catalase increases the auto-oxidation of Aminochrome o-semiquinone. However, in the presence of DT-diaphorase, as it is shown in the right, Aminochrome is reduced to Aminochrome o-hydroquinone. Although o-hydroquinones are also auto-oxidized; the rate of auto-oxidation is lower than that of aminochrome o-semiquinone. One of the possibilities is that SOD and Catalase recognize a general motif in the structure of the o-hydroquinone and prevents the generation of the degenerative o-semiquinones. Interestingly, the alcohols are present in the benzene rings of dopamine and o-hydroquinone. The cyclilization of the amine group is the only feature that differs among dopamine and aminochrome o-hydroquinone. The hydroquinone is a stable structure that is depleted from cells by reactions mediated by sulfo-transferases and COMT.
om superoxide species, thus moving the reaction towards the reactant (aminochrome). Thus, the presence of SOD and catalase increases the auto-oxidation of Aminochrome o-semiquinone. However, in the presence of DT-diaphorase, as it is shown in the right, Aminochrome is reduced to Aminochrome o-hydroquinone. Although o-hydroquinones are also auto-oxidized; the rate of auto-oxidation is lower than that of aminochrome o-semiquinone. One of the possibilities is that SOD and Catalase recognize a general motif in the structure of the o-hydroquinone and prevents the generation of the degenerative o-semiquinones. Interestingly, the alcohols are present in the benzene rings of dopamine and o-hydroquinone. The cyclilization of the amine group is the only feature that differs among dopamine and aminochrome o-hydroquinone. The hydroquinone is a stable structure that is depleted from cells by reactions mediated by sulfo-transferases and COMT.
Question: How pertinent is the expression of DT-diaphorase for the progression of parkinson's disease, do these patients have a lower expression of DT-diaphorase in the substantia nigra? Do dopaminergic cells that express high levels of DT-diaphorase in the brain less likely to die from the disease?

